




© С. Дробышевский, 2017
© Р. Евсеев, иллюстрации, 2017
© Е. Мартыненко, иллюстрации, 2017
© О. Федорчук, иллюстрации, 2017
© А. Бондаренко, оформление, 2017
© ООО “Издательство АСТ”, 2017
Издательство CORPUS ®
* * *
Посвящается Инге, Володе и Маше –
моей любимой семье
…а также муравьеду и руконожке
Рассказ о цепях коротких и длинных, непрерывных и оборванных, магистральных и параллельных, прямых и извилистых, о том, из каких звеньев они составлены, и о том, все ли звенья достают, как люди достают эти звенья и как звенья достают людей, о звеньях прочных и не очень, о звеньях главных и второстепенных, о звеньях между звеньями и рядом со звеньями, неотвратимости и случайности, о наследии и следах, много о прошлом, в меру о настоящем и немножко о будущем…
Люди
Часть десятая, последняя, из которой Читатель узнает всю правду о своем происхождении, двуногой родне – дальней и ближней, пращурах, похожих на обезьян и на людей, предках прямых, двоюродных и троюродных, пра1000-дедушках и пра1000-бабушках, их подвигах и неудачах, стремлениях и успехах
Пролог
Внепрерывной череде эволюционных событий нет знаков препинания. Родители и потомки почти не отличаются друг от друга, нужны миллионы лет, чтобы количество перешло в качество, и лишь на огромных временных отрезках мы замечаем появление чего-то совсем нового. Поэтому невозможно сказать, в какой конкретно момент начинается “совсем-совсем” наша линия эволюции, когда “совсем” обезьяны становятся “совсем” людьми. Процесс занял сотни тысяч поколений. Изменения климата и колебания уровня океанов, конкуренты и хищники, соседи и унаследованные от предков свойства – причудливая смесь причин и побуждений двигала эволюцию. По пути возникали причудливые существа, многие из них имели неплохие шансы стать истинно разумными, а некоторые даже и стали. Итог известен – мы остались единственным светочем интеллекта на планете. А что привело нас к этому?..
Гоминиды Hominidae включают австралопитековых Australopithecinae и собственно людей Homininae – и пусть кладисты проклянут меня за то, что я отделяю их от шимпанзе и горилл!
Границы подсемейств австралопитековых и людей, как обычно, не слишком четкие. И проблема здесь не в нашей неосведомленности, а как раз в избытке знаний: в последовательном ряду плавно меняющихся форм трудно провести четкую границу: досюда – обезьяны, тут они кончаются, а здесь вот начинаются люди.
Уголок занудства
Для того чтобы избежать опасной конкретики, иногда применяются стадиальные названия. Австралопитеков изредка называют протантропами. Около 2 млн лет назад их потомки потеряли практически все специфические черты сходства с обезьянами и могут называться первыми настоящими людьми, или гомининами; иногда, чтобы подчеркнуть их “истинность”, пользуются термином “эугоминины” – это все люди от хабилисов до нас. Эугоминины, в свою очередь, разделяются на последовательные стадии. Homo habilis и близких гоминид часто называют “ранними Homo” или, реже, эогомининами (“эу” – настоящий, а “эо” – ранний). Из них возникли архантропы (ранних – Homo ergaster и подобных им – можно называть преархантропами, а поздних – Homo heidelbergensis – препалеоантропами). За ними следуют палеоантропы, а венчают процесс неоантропы.
До середины XX века считалось, что стадии в процессе накопления эволюции трансформировались и плавно или же скачкообразно сменяли одна другую. В каждый момент времени на Земле существовали представители лишь одной стадии. Такая концепция получила название стадиальной теории. Она подкреплялась также представлениями о последовательной смене археологических культур: олдувайской на ашельскую (нижний палеолит), последней – на мустьерскую (средний палеолит), которую заместил верхний палеолит. Носителями каждой культуры была соответствующая стадия гоминин.
В настоящее время практически все ученые склоняются к мысли о более сложном характере эволюции. Дело даже не в том, что некоторые популяции обгоняли остальных по уровню своего развития, а в том, что они эволюционировали по-разному. Кроме того, миграции или же изоляция играли значительную роль в неравномерности распространения признаков по планете. Отдельные особенности могли возникать в разных группах независимо, объединяясь потом вследствие смешений. Такой вариант эволюции называется сетевидной эволюцией.
Стадиальные термины до сих пор применяются в научной литературе, но только для удобства разделения материала. В реальности слова “архантропы”, “палеоантропы” и “неоантропы” объединяют подчас весьма отличающиеся группы гоминин. Преимуществом их использования является уход от запутанной систематической номенклатуры, понимаемой разными исследователями по-разному.
Обычно древнейшей границей гоминид считают момент отделения ветви шимпанзе. Проблема в том, что ископаемых шимпанзе-то как раз у нас и нет. Вообще палеонтологическая летопись в Африке между 14 и 6 млн лет до обидного неполна – это большей частью изолированные зубы (в Азии, напротив, из этого интервала полно сивапитеков, но они мало что могут рассказать о нашей эволюции). Пока не найдены богатые окаменелостями отложения этого немаленького интервала, остается довольствоваться останками самбуропитека, отавипитека, неназванных гоминоидов Туген-Хиллс и Накали, хорорапитека и накалипитека. Как-никак, а все же они заполняют таинственный промежуток.
Отсутствие богатых находок тем досаднее, что это время было одним из самых важных для нашего появления: гоминоиды выходили в саванны и становились прямоходящими. Пионеры открытых пространств еще не слишком отличались от своих предков, да и не выскочили они сразу на равнину. Долгие и долгие миллионы лет человекообразные жили в редколесьях – леса не высохли в один момент, замещение экосистем происходило достаточно долго, чтобы приматы смогли адаптироваться к новым условиям.
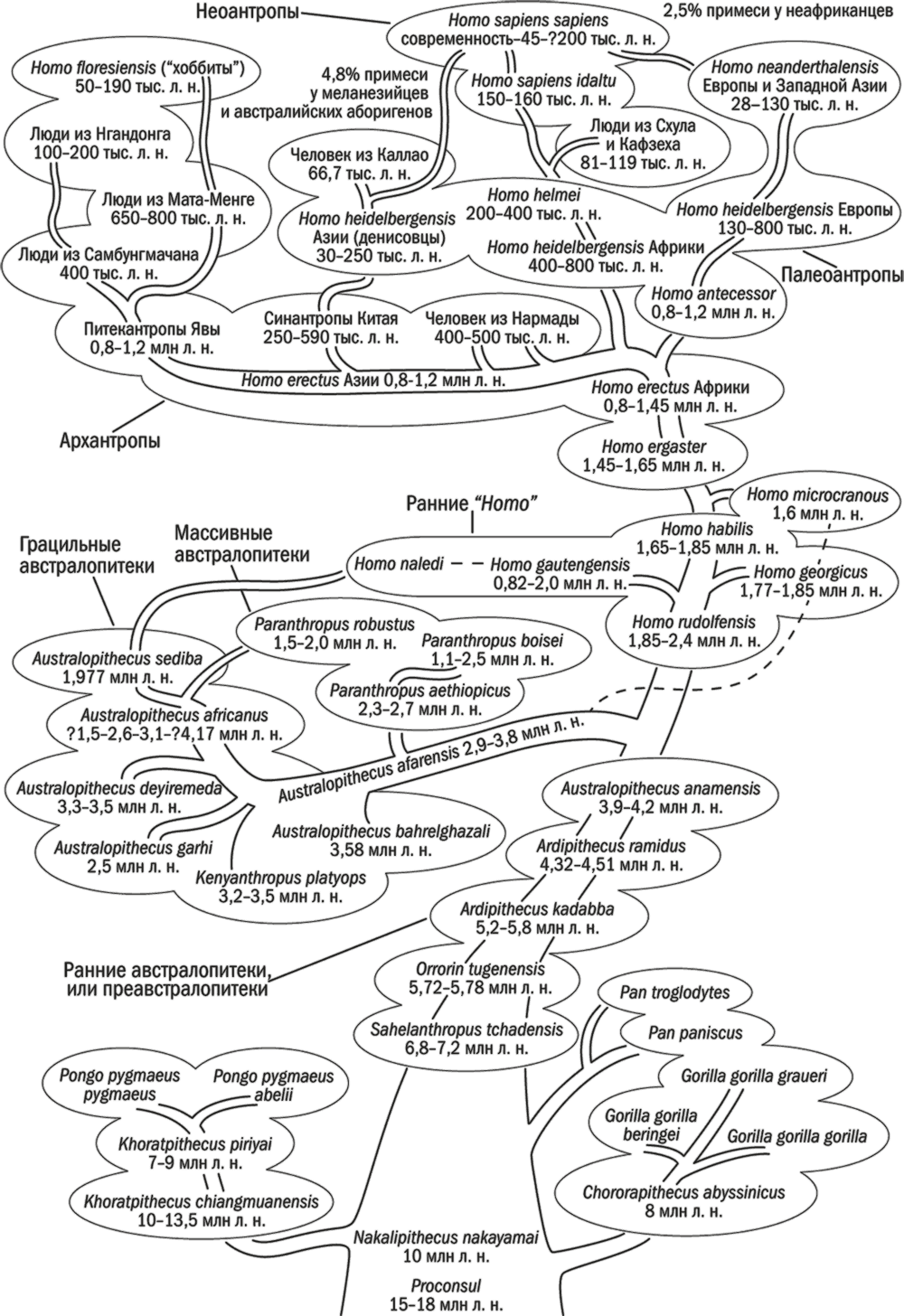
Рис. 1. Схема эволюции гоминид.
Поэтому первые почти-прямоходящие и прямоходящие в строении зубов и черепа имеют намного больше обезьяньего, чем человеческого, и потому выделяются в самостоятельное подсемейство австралопитековых Australopithecinae. Правда, самые ранние из них, открытые сравнительно недавно, отличаются от тех, которых находили на протяжении XX века, отчего многие антропологи остерегаются называть их австралопитеками, предпочитая термин “преавстралопитеки”. Все же удобнее разделять австралопитековых на три группы: ранних, грацильных и массивных.
Австралопитеки – ключевая группа эволюции гоминид. Насколько известно, они никогда не покидали пределов Африки. Находки вне Черного континента, в прошлом приписывавшиеся австралопитекам (Убедийя из Израиля, Мегантроп 1941 года и Моджокерто с Явы), либо крайне фрагментарны и, следовательно, спорны, либо при ближайшем рассмотрении оказались эректусами. Также в Агригенте на Сицилии была найдена загадочная верхняя челюсть, датированная 3,5–5,0 млн лет назад и описанная в 1985 году Дж. Бианчини как Australopithecus sicilianus, но она, судя по всему, принадлежала какой-то обезьяне.
Внутри Африки местонахождения австралопитеков концентрируются в двух основных районах: Восточной Африке (Танзания, Кения и Эфиопия) и Южной Африке. Отдельные находки были сделаны также в Северной Африке; думается, их малое количество связано больше с условиями захоронения и малой изученностью региона, а не с реальным расселением австралопитеков. Ясно, что в столь широких временных и географических рамках природные условия не раз менялись, что приводило к появлению новых видов и родов.
Одновременное сосуществование разных “хороших” видов австралопитеков на одной территории не было твердо доказано ни для одного местонахождения, хотя предположений на этот счет выдвигалось много. Однако сосуществование поздних австралопитеков с “ранними Homo” не вызывает сомнения.
Образ жизни австралопитеков и “ранних Homo”, видимо, был не похож на известный у современных приматов. Они жили в широком диапазоне условий, но чаще в той или иной степени облесенных саваннах. Вообще, реконструированные африканские ландшафты времен первых гоминид чаще всего чрезвычайно мозаичны и часто на небольшой территории включают в себя элементы от открытых сухих саванн до влажных тропических лесов. Возможно, разнообразие ландшафтных зон препятствовало крайней специализации австралопитеков; истоки высокой экологической пластичности современного человека, видимо, можно искать уже в том времени. Австралопитеки питались преимущественно растениями, но гоминиды с самого появления были так или иначе всеядны; мы точно знаем, что поздние австралопитеки и первые Homo охотились на антилоп или отнимали добычу у крупных хищников – львов и гиен.
Некоторые линии австралопитеков зашли в эволюционный тупик. Но они были той единственной уникальной группой, которая смогла перешагнуть грань животного состояния и ступить на путь очеловечивания. И сейчас, благодаря усердному труду антропологов, мы знаем основные этапы и даже многие детали этого пути. А “недостающее звено” превратилось в длинную череду “достающих”.
Глава 1
Обезьянолюди: ранние австралопитеки
Ранние австралопитеки населяли Африку между 7 и 3,9 млн лет назад. В конце миоцена леса исчезали, саванны наступали, и вот под барабанную дробь, звонкие литавры и голосистые фанфары на сцену выходит…
Сахелянтроп Sahelanthropus tchadensis! И не беда, что у него голова с кулак, ничего, что больше всего он похож на карликовую гориллу, – это действительно он, Великий Предок!
Череп и нижняя челюсть сахелянтропа были найдены французскими исследователями в Северной Африке, в Республике Чад, в Торос-Меналла, и описаны в 2002 году (Brunet et al., 2002); позже добавились еще две нижние челюсти и новый зуб (Brunet et al., 2005). Это древнейший и примитивнейший представитель нашей родной эволюционной линии. По фауне отложения с останками сахелянтропа датированы 6–7 млн лет назад; к сожалению, фауна не меняется быстро, а вулканов, по пеплу коих можно было сделать точную датировку, поблизости не оказалось, так что поначалу приходилось довольствоваться погрешностью в миллион лет. Однако прогресс идет, и уже скоро по соотношению изотопов бериллия в костях антракотериев была получена на полмиллиона лет более точная дата – 6,8–7,2 млн лет назад (Lebatard et al., 2008).

Рис. 2. Череп Sahelanthropus tchadensis.
Череп получил коллекционный номер TM 266-01-60-1, но этот скучный паровозный код, конечно, не мог в должной мере отразить ликование антропологов, которые по сложившейся традиции окрестили его личным именем Тумай. Нам безмерно повезло, что от древнейшего гоминида сохранился целый череп, хотя бы и сильно перекошенный; деформацию же выправили трудолюбивые реставраторы (Zollikofer et al., 2005).
Кстати, об именах…
Антропологи тоже любят украшать себе жизнь. Так среди австралопитеков и “ранних Homo” появились “Беби из Таунга”, “Люси”, “Сын Люси”, целых два “ребенка Люси” (одна из них – “Дочка Люси”, или “Селам”), “Первое семейство”, “Миссис Плез” и “Мистер Плез”, “Тумай” и “Арди”, “Миллениум Мен”, или “Человек Тысячелетия”, “Кадануумуу”, или “Большой Человек”, “Загадочный череп” и “Черный череп” (он же “череп Дарта Вейдера”), “Абель”, “Джордж” и “малыш Джонни”, “Твигги”, “Синди”, “Орфей” и “Эвридика”, “Щелкунчик” и даже такие почти индейские имена, как “Колено Джохансона” и “Маленькая Стопа” (он же “Синдерелла”).
Череп Тумая совсем маленький, с объемом мозга всего 360–370 см³ – меньше, чем в среднем у обыкновенного шимпанзе, но чуть больше, чем у бонобо. Лоб невероятно плоский, череп сверху почти совсем ровный: по этому признаку сахелянтроп выглядит примитивнее любой человекообразной обезьяны. Получается так потому, что лицо сахелянтропа расположено впереди от мозговой коробки, а не под ней, как это обычно для человекообразных. Рельеф черепа не по размеру мощный: надбровье выступает хотя и не столь резко, как у шимпанзе, но фактически рекордно для австралопитековых; височные линии ближе к затылку сливаются в сагиттальный гребень; выйный гребень так вообще выглядит как широкая пластина, идущая от височных костей поперек затылка, заостренная по краю и загнутая вниз. Рельефом сахелянтроп однозначно превосходит шимпанзе, что загадочно. Возможно несколько объяснений. Во-первых, сахелянтроп мог питаться гораздо более жесткой пищей. Во-вторых, переход к прямохождению мог сопровождаться усилением шейной мускулатуры, так как голова держалась на позвоночнике наискосок, ее надо было уравновешивать сзади, рычагом для чего и служил выдающийся затылочный гребень.
Одно из самых замечательных свойств сахелянтропа – положение большого затылочного отверстия. Как уже говорилось, у четвероногих обезьян оно расположено сзади и ориентировано назад, а у человека – сдвинуто на середину основания черепа и смотрит вниз. У сахелянтропа же его положение и ориентация строго промежуточны. Видимо, это существо проводило на двух ногах гораздо больше времени, чем шимпанзе, но и совсем прямоходящим его назвать трудно. Вообще, основание черепа – самое продвинутое место сахелянтропа.
Лицо Тумая относительно мозгового отдела заметно уменьшено. При взгляде на него в душе истинного антрополога распускаются ромашки. Ведь именно таким положено быть лицу настоящего Великого Предка: в меру крупное, не уплощенное в горизонтальной плоскости и выступающее вперед в вертикальной, но без крайностей, характеризующих горилл и шимпанзе. Анфас Тумай весьма напоминает некоторых самок бонобо: глазницы квадратные, нос широкий, альвеолярный отросток умеренно высокий. Однако по ряду признаков сахелянтроп отличается от шимпанзе: так, у него очень широкое межглазничное расстояние. В профиль же он ни на кого не похож, разве что на ардипитека: верхняя половина лица вертикальна, а челюсти равномерно выдвинуты вперед. Даже от грацильных австралопитеков отличия достаточно очевидны: у Тумая намного слабее развит альвеолярный прогнатизм (Guy et al., 2005). Правда, нижняя челюсть выглядит весьма тяжелой, даром что она невелика.
Зубы сахелянтропа довольно крупные, но их относительные пропорции замечательны: тогда как моляры больше, чем у шимпанзе, клыки – намного меньше. И это притом, что Тумай – самец! Тут может быть два объяснения: первым делом так и подмывает сказать, что клыки сахелянтропа редуцировались и это весьма прогрессивно; однако, учитывая данные по проконсулам, приходится признать, что скорее уж правильнее говорить, что это у шимпанзе клыки увеличились. Если взять по модулю уменьшение от проконсула к сахелянтропу и увеличение от него же к шимпанзе, то шимпанзе оказывается самым прогрессивным, коли уж прогрессом мы считаем отличие от исходника.
Судя по составу фауны, местность, где жил сахелянтроп, включала и водоемы, и саванны, и леса (Le Fur et al., 2014), так что у него был широкий выбор, где жить и чего есть.
Конечно, наука на то и наука, что дело не могло обойтись без скептиков, подбросивших ложку дегтя в праздничное варенье первооткрывателей Тумая (Wolpoff et al., 2006). Они указали на целый ворох примитивностей сахелянтропа, роднящих его с миоценовыми человекообразными типа кениапитека, уранопитека и даже гигантопитека. По ряду параметров основания черепа Тумай больше схож с некоторыми самками горилл, нежели австралопитеками, не говоря уж о людях. Однако чего и ждать от Великого Предка? На то он и предок, чтобы быть похожим на других родственников.
В итоге многие ученые склонны отводить сахелянтропу почетную роль “двоюродного прапрапрадедушки”, но не считают его одним из наших непосредственных предков. Для окончательного прояснения вопроса нужны новые находки. Пока их явно недостаточно, но богатство местонахождения Торос-Меналла оставляет надежду на скорые открытия.
К великому сожалению, от сахелянтропа не сохранились кости посткраниального скелета. Зато они найдены в Кении. Из ее земли на эволюционную арену на почти прямых ногах ковыляет…
Оррорин Orrorin tugenensis. Его останки фрагментарнее, но одновременно многочисленнее. Их обнаружили также французские антропологи в местности Туген-Хиллс на исходе 2000 года, отчего новооткрытый Великий Предок получил по-голливудски броское прозвище Миллениум Мен – Человек Тысячелетия. Официальное описание состоялось в 2001 году. (Senut et al., 2001). Кости были найдены в области Баринго, в местности Капчеберек, в формации Лукейно, и были датированы 5,72–5,88 млн лет назад (Deino et al., 2002).
Дюжина окаменелостей включала два куска нижней челюсти, зубы, фрагменты плечевой, фаланги кисти, правой и двух левых бедренных костей. В этом суповом наборе поместилось аж пять индивидов, как самцов, так и самок. Следы зубов кошачьего на бедренной кости повествуют печальную историю о нелегкой судьбе пионеров прямохождения…
Кстати, о дежавю и пользе статистики…
На самом деле, останки орроринов были известны и раньше, только тогда никто не знал, что это и с чем его надо есть. Еще в 1974 г. в Кении, в Лукейно, в местонахождении Чебойт, был найден фрагмент челюсти с нижним моляром KNM LU 335, но долгие годы – почти тридцать лет – он оставался “вещью в себе”, непонятным добавлением к существовавшей тогда картине эволюции австралопитеков, так как его было не с чем сравнить. Было ясно, что он очень древний (5,72–5,88 млн лет назад), похожий и на шимпанзе, и на человека, сравнительно небольшой, но лишь после описания более представительных материалов он обрел смысл.
Зубы орроринов размером примерно как у шимпанзе, более поздних ардипитеков и афарских австралопитеков, но клыки сравнительно небольшие, особенно в сравнении с шимпанзе.
Гораздо больше информации можно извлечь из длинных костей оррорина. Обломок плечевой кости очень мощный, очевидно, руки у оррорина были могучие, судя по специфике прикрепления мышц – отлично приспособленные к лазанию по деревьям. О том же свидетельствует сильный изгиб фаланги кисти.
Главный интерес вызвали бедренные кости. Одной из важнейших особенностей оррорина оказался хорошо выраженный комплекс прямохождения. Сравнение их с костями современных человекообразных обезьян и людей позволило утверждать, что уже почти 6 млн лет назад в Восточной Африке жили практически полностью прямоходящие существа. Самое поразительное – по некоторым признакам оррорины выглядят даже более прогрессивными, чем гораздо более поздние афарские австралопитеки (Pickford et al., 2002). Некоторые журналисты даже переврали это в обычно-сенсационном ключе, объявив о “перевороте в представлениях о человеческой родословной”: дескать, вовсе не известные доныне австралопитеки были нашими действительными предками, а новооткрытые оррорины. Особо обращает на себя внимание прямизна бедренной кости, передне-задняя уплощенность, длина и наклон ее шейки, а также сферичность головки и другие тонкости морфологии. Впрочем, другие исследователи более осторожно подходят к вопросу (Richmond et Jungers, 2008).
Оррорины были совсем некрупными созданиями – около 1,1–1,2 м (максимум 1,44) ростом и 35–50 кг весом (Nakatsukasa et al., 2007). Видимо, это был средний размер для всех ранних и большинства грацильных австралопитеков.
Скачут ножки по дорожке: такие ли уж прыткие были оррорины?
Многое в жизни зависит от точки зрения. В палеоантропологии дело часто обстоит так же. Потому-то одни и те же находки разные исследователи склонны расценивать совершенно по-разному. Кости не меняются, а их интерпретации разнятся до противоположностей. Не избежал этой чехарды мнений и Orrorin tugenensis – один из древнейших известных гоминид.
Находка была разрекламирована как сенсация века и даже тысячелетия. Шутка ли – получалось, что именно Человек Тысячелетия был нашим прямым предком, а ставшие уже привычными афарские австралопитеки – просто тупиковая ветвь.
Однако в первых описаниях бедренные кости орроринов сравнивались только с современными видами. А ведь нынешние человекообразные обезьяны – вовсе не те звери, что скакали по ветвям миоценовых лесов миллионы лет назад. И гориллы, и шимпанзе прошли собственный долгий путь эволюции, особенности их бедренных костей, хоть частенько и считаются примитивными, в действительности сформировались едва ли не позже нашего прямохождения и имеют специфику, в немалой степени обусловленную адаптацией к полуназемному образу жизни. Исправить этот пробел сравнительной палеоантропологии взялась международная испано-американо-итальянская команда антропологов (Almecija et al., 2013). Они сравнили бедренные кости орроринов не только с современными обезьянами и людьми, но и миоценовыми испанопитеком, дриопитеком, экваториусом и проконсулом. Из этой плеяды только проконсул является вероятным предком орроринов, но эволюционный уровень у них всех заведомо примитивный. Результат сопоставления оказался полностью закономерным, хотя для кого-то, может быть, и неожиданным.
Во-первых, современные орангутаны, гориллы и шимпанзе отличаются от миоценовых гоминоидов, причем по некоторым чертам в ту же сторону, что и современный человек; гиббон, что характерно, сохранил древнее строение. Во-вторых, по тем же признакам оррорин оказывается примитивным, схожим с приматами, жившими 15–20 млн лет назад, но в целом промежуточным между миоценовыми обезьянами и более поздними австралопитеками, от которых те же признаки плавно видоизменяются до состояния “ранних Homo” и далее до современного человека.
Самой показательной чертой оказалось расположение ягодичной бугристости: у миоценовых гоминоидов, современных гиббонов и оррорина она смещена на боковую поверхность кости, а у современных крупных человекообразных обезьян, древних гоминид и людей расположена на задней стороне бедра. А ведь на эту бугристость крепится ягодичная мышца, которая у разных существ может выполнять удивительно разнообразные функции. У человека она выпрямляет ногу, тогда как у обезьян скорее отводит ее в сторону. У орроринов она, получается, была совсем дремучей, а нога не могла полностью распрямляться в тазобедренном суставе. Столь же архаично выглядят и другие признаки Человека Тысячелетия: морфология головки бедра, угол шейки к диафизу, выступание вбок основания большого вертела и завернутость внутрь малого вертела.
Таким образом, миф о необычайной продвинутости прямохождения оррорина не подтвердился. Конечно, это не отменяет наличия у оррорина действительно очень прогрессивных черт, например очень длинной шейки бедра, уплощенной спереди назад, а также сферической формы головки.
Получается, что Orrorin tugenensis закономерно занимает промежуточное положение между древнейшими человекообразными и современными людьми, тогда как нынешние крупные человекообразные вовсе не копии своих ископаемых предков. Только гиббоны, чьи пращуры никогда не спускались с деревьев, сохранили примитивное состояние бедренной кости. Зато как поменялись их руки!
В очередной раз можно убедиться, что все течет, все меняется, эволюция – постоянный процесс и никакие биологические объекты не замирают в морфологическом трансе (ну, разве кроме щитней Triops cancriformis, у которых День сурка начался 230 млн лет назад да так и не закончился, но это совсем-совсем другая история…).
Таким образом, оррорины, видимо, вставали на две ноги, но четвероного-древесное прошлое еще явно сквозило в их облике. Время меж тем шло, прогресс тоже не стоял на месте. Появлялись новые виды…
Ардипитек кадабба Ardipithecus kadabba – пока недостаточно изученное, но очень важное звено эволюции человека (Haile-Selassie, 2001). Его разрушенные временем останки были найдены в Эфиопии, в долине реки Средний Аваш, в местонахождениях Алайла и Аса-Кома 3; они имеют возраст 5,2–5,8 млн лет назад. Первые находки были сделаны еще в 1997 году, но продолжались и позже: в 2002 году там же (Haile-Selassie et al., 2004), а после похожие зубы были найдены в Гоне (Simpson et al., 2007).
От кадабб сохранились в основном зубы, но есть обломки нижней челюсти, ключицы, двух плечевых и локтевой, две фаланги кисти и одна – стопы. Не будучи Ж. Кювье, не так легко представить облик и образ жизни существ по столь скудным останкам. Все же ясно, что кадаббы были мелкими созданиями: нижняя челюсть меньше, чем у афарских австралопитеков, хотя и массивнее, чем у них. Клыки кадабб сопоставимы с клыками мелких самок шимпанзе: круглые в сечении, конические по форме, они превосходят клыки орроринов и более поздних гоминид. Первые нижние премоляры, найденные в Среднем Аваше, секториальные: это значит, что их коронка асимметрично скошена, чтобы большому верхнему клыку было куда поместиться. Любопытно, что в Гоне этот же зуб не секториальный – видимо, некоторые популяции были прогрессивнее.
Ключица, плечевые и локтевая имеют весьма небольшие размеры, но выраженный рельеф, свидетельствующий о неплохих способностях к лазанию по деревьям. Фаланги кисти принципиально не отличаются от таковых афарских австралопитеков, то есть сочетают обезьяньи и человеческие признаки примерно поровну.
Особенно интересна проксимальная фаланга стопы: с одной стороны, она сильно изогнута, как у человекообразных обезьян, с другой – суставная поверхность ее основания ориентирована вверх, как то характерно для афаренсисов и современных людей, что косвенно свидетельствует о прямохождении.
Кстати, опять о статистике…
Еще в 1967 г. в Лотагаме в Кении был найден фрагмент нижней челюсти с одним моляром KNM – LT 329, имеющий древность 5,0–5,5 млн лет назад. Он долго оставался “бедным родственником” – безымянным и не находящим себе места в общей родословной, а ныне отнесен к ардипитекам.
Все вышеназванные находки интересны и замечательны, но меркнут на фоне Великого Предка – ардипитека рамидуса.
Ардипитек Рамидус Ardipithecus ramidus – образцовый обезьяночеловек, то самое достающее звено, которое гипотетически реконструировали еще в XIX веке, потом так долго искали и так жаждали увидеть воочию. В нем воплотились все научные предсказания, сделанные антропологами за полторы сотни лет. Более того – он превзошел все ожидания и возложенные на него надежды!
Кстати, еще разок о статистике…
Надо сказать, что останки ардипитеков были известны задолго до описания в качестве самостоятельного вида. Еще в 1980-х годах в кенийском местонахождении Табарин, в формации Чемерон, были найдены обломок нижней челюсти с двумя молярами KNM-TH 13150 (4,43 млн лет назад) и кусок плечевой кости KNM-BC 1745 (5,1 млн лет назад). К кому их только не относили! И только с открытием многочисленных находок ардипитеков мы теперь можем уверенно говорить, что в Табарине жили представители этого вида.
В 1993 году в эфиопской местности Арамис в долине реки Средний Аваш антропологи нашли россыпь обломков. В 1994 году по ним был описан вид Australopithecus ramidus (White et al., 1994), а спустя год он был переименован в Ardipithecus ramidus (White et al., 1995). Их возраст – 4,4 млн лет. После фрагменты челюстей, зубы и фаланги этого же вида обнаружились в эфиопском местонахождении Аз-Дума в местности Гона (4,32–4,51 млн лет назад). Показательно, что там же в более древних слоях встречены останки A. kadabba – наглядная преемственность, напрашивающаяся в учебники.
История изучения скелета ардипитека – ярчайший пример научной добросовестности. Ведь между его обнаружением – 29 декабря 1993 года – и полноценной публикацией – 2 октября 2009 года – прошло почти 16 лет! Об уникальной находке было заявлено сразу же, но подробности все это время оставались неизвестны, так что у некоторых исследователей даже зародилось сомнение: “а был ли мальчик”? Все эти долгие годы международная группа исследователей, в том числе первооткрыватель – Иоганнес Хайле-Селассие, трудились над сохранением рассыпающихся костей, реконструкцией раздавленного в бесформенный комок черепа, описанием морфологических особенностей и поиском функциональной интерпретации мельчайших подробностей строения костей. Ученые не пошли по пути предъявления миру очередной скороспелой сенсации, но действительно глубоко и тщательно исследовали самые разные аспекты находки. Для этого пришлось изучить такие тонкости сравнительной анатомии современных человекообразных обезьян и человека, которые до сих пор оставались неизвестными. Естественно, к сравнению были привлечены и данные по множеству ископаемых приматов и австралопитеков. Мало того, детальнейшим образом были рассмотрены геологические условия захоронения ископаемых останков, древняя флора и фауна, что позволило реконструировать среду обитания ардипитеков достовернее, нежели для многих более поздних австралопитеков.
В итоге все части скелета ардипитека были подробнейшим образом описаны в серии статей: череп (Suwa et al., 2009a), зубы (Suwa et al., 2009b), рука и кисть (Lovejoy et al., 2009b), таз и нога (Lovejoy et al., 2009d), стопа (Lovejoy et al., 2009a), на основании чего были сделаны выводы о его способе передвижения (Lovejoy et al., 2009с) и путях возникновения бипедии в целом (Lovejoy, 2009).
Новоописанный скелет ардипитека являет собой замечательный пример подтверждения научной гипотезы. В своем облике он идеально сочетает признаки обезьяны и человека. Фактически тот образ, который полтора столетия будоражил воображение антропологов и всех, кому небезразлично наше происхождение, стал наконец реальностью. И все равно, сколь тщательно антропологи ни вымысливали и ни высчитывали стати Великого Предка, ардипитек удивил всех.
Находки в Арамисе многочисленны – останки принадлежат не менее чем 21 особи, но наиболее важен скелет взрослой самки ARA-VP-6/500, от которого осталось около 45 % костей (больше, чем от знаменитой Люси – самки афарского австралопитека из Хадара с древностью 3,2 млн лет назад!), в том числе почти весь череп, хотя и в крайне деформированном состоянии. “Арди” – так нежно прозвали Великую Прародительницу – имела рост около 1,2 м и могла весить до 50 кг. Существенно, что половой диморфизм ардипитеков был выражен гораздо слабее, чем у шимпанзе и даже более поздних австралопитеков, то есть самцы были ненамного крупнее самок; это мы знаем благодаря сравнению клыков разных особей. Объем мозга Арди достигал 300–350 см³ – столько же, как у сахелянтропа, но меньше, чем обычно у шимпанзе.
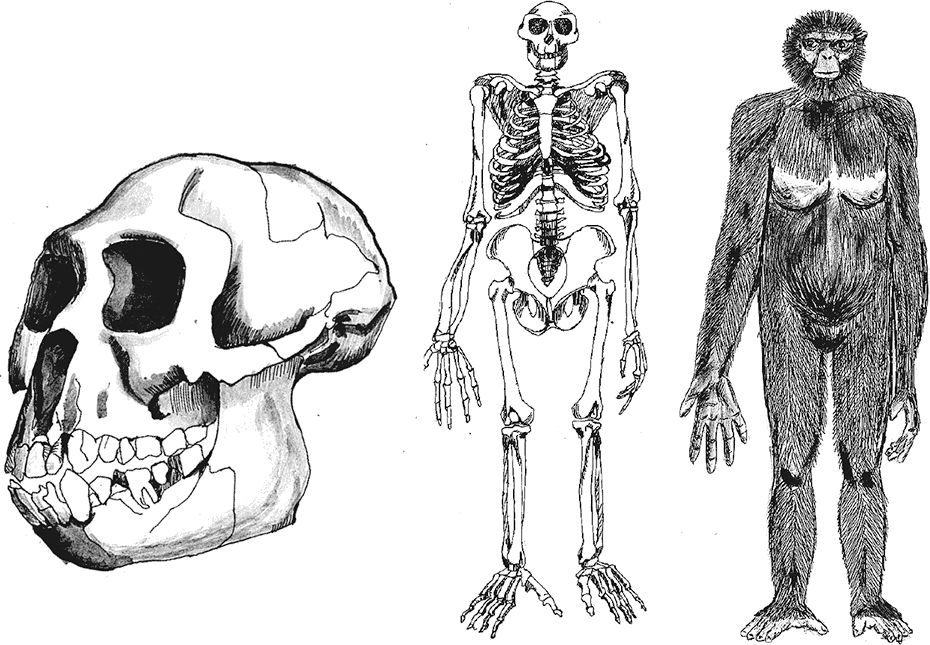
Рис. 3. Череп, скелет и реконструкция Ardipithecus ramidus.
Строение черепа ардипитека довольно примитивно, фактически обезьянье. Арди была бы мало отлична от мелкой самки шимпанзе, если бы не маленькие лицо и клыки и не сдвинутое вперед затылочное отверстие. Череп очень мал, со сравнительно слабым рельефом, без сагиттального гребня. Лоб более выпуклый, чем у сахелянтропа, маленькое слуховое отверстие и нижнечелюстная ямка височной кости – важные диагностические признаки – напрочь обезьяньи. Глазницы большие, межглазничное пространство не слишком широкое, нос можно даже назвать узким. Лицо относительно мозгового отдела довольно маленькое, но челюсти сильно выступают вперед, скуловые дуги широко расставлены, а между верхним вторым резцом и клыком имеется небольшая диастема – промежуток для нижнего клыка. Нижние челюсти ардипитеков не слишком велики, в целом как у шимпанзе и австралопитеков. Зубы имеют строение, промежуточное между шимпанзе и афарскими австралопитеками, причем по некоторым чертам ближе именно к шимпанзе. Уникальной особенностью рамидусов оказалась преимущественная редукция верхних, а не нижних клыков, так что последние оказались крупнее и обезьяноподобнее первых, больше выступающими за ряд прочих зубов.
- Происхождение жизни. От туманности до клетки
- Достающее звено. Книга 1. Обезьяны и все-все-все
- Достающее звено. Книга 2. Люди
- От атомов к древу: Введение в современную науку о жизни
- #Панталоныфракжилет. Что такое языковые заимствования и как они работают
- Против часовой стрелки. Что такое старение и как с ним бороться
- Евангелие от LUCA. В поисках родословной животного мира
- Археология русского интернета. Телепатия, телемосты и другие техноутопии холодной войны
- Достающее звено. Книга 1. Обезьяны и все-все-все
- Достающее звено. Книга 2. Люди